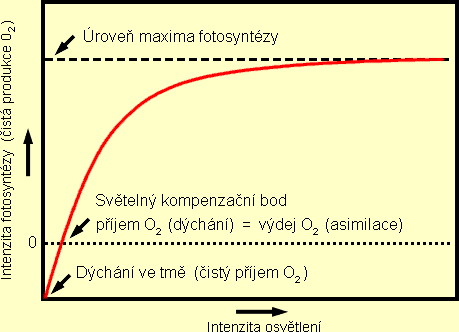
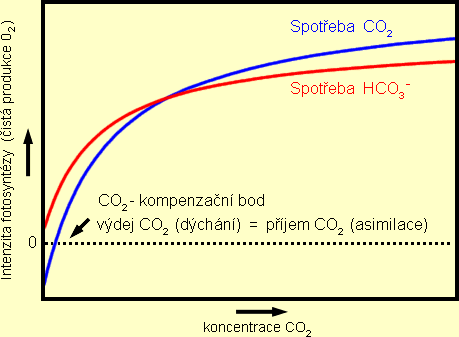
Nízká dostupnost CO2 je ve vodním prostředí způsobena zejména
nízkou (difuzní) rychlostí pronikání molekul CO2 vodním sloupcem. (Podrobněji je
problematika rozebrána v textu
plyny ve vodě - difuze
a výživa rostlin.) Může však nastat - a zcela běžně nastává
- stav, kdy rostliny mají ve vodě sice dostatek rozpuštěného CO2 ,
ale pro nedostatek světla nemůže být plně využit při fotosyntéze. Tento stav je obvyklý
ve stojatých vodách, kde v hlubších vrstvách může být vysoká koncentrace CO2
(způsobená např. mineralizací organických usazenin na dně), avšak rostliny nemohou růst do
větší hloubky kvůli nízké intenzitě záření pronikajícího sloupcem vody.
Rostliny mají vyvinutu důležitou schopnost regulovat množství enzymů (např. Rubisco,
PEP-karboxyláza) účastnících se fixace uhlíku. Zvyšuje-li dostatek CO2 účinnost
využití světla, pak umožňuje submerzním rostlinám pronikat i do hloubky, ve které je již
intenzita světla (FAR) velmi nízká.
V intenzívně osvětlených porostech vodních rostlin (např. v mělkých stojatých vodách
s dostatkem světla) může naopak dojít k rychlému odčerpání oxidu uhličitého z prostředí
a jeho deficit znemožní rychlý růst rostlin. Při nízké koncentraci CO2 mohou
rostliny vytvořit více enzymů důležitých pro vázání CO2 při fotosyntéze (zlepší
se jeho "čerpání" z nízkých koncentrací v okolní vodě) a dokáží tím zvýšit využití
dostupné energie světla pro růst rostlin. Vysoká dostupnost světla tak umožní submerzním
rostlinám snížit úroveň kompenzačního bodu CO2 , což neznamená nic jiného, než
schopnost růstu rostliny při nižší (mezní) koncentraci CO2 ve vodě. Tato
adaptační funkce ponořených vodních rostlin na deficit obou faktorů je však velmi
náročná na spotřebu energie v porovnání se stavem dostatečného přísunu světla i CO2 .
Podobně je možno přistupovat např. k ovlivňování intenzity fotosyntézy teplotou a současně
i koncentrací CO2 (za předpokladu nasycení ostatními minerálními živinami a dostatkem
světla) - při zvýšené teplotě se zvýší produkce zelené hmoty až do stavu nasycení, ale další
zvyšování teploty vede k poklesu tvorby biomasy. Současným zvýšením koncentrace CO2
se urychlí i produkce biomasy (křivka se posune k vyšším hodnotám intenzity fotosyntézy).
Množství různých interakcí (vzájemného ovlivňování) jednotlivých růstových faktorů je
příliš rozsáhlé, současná věda (rostlinná fyziologie) nedokáže dosud všechny vzájemné
souvislosti analyzovat - výzkumy jsou značně obtížné a nákladné. Řada výzkumů (zaměřených
především na vlivy vzrůstající koncentrace CO2 v atmosféře na kulturní plodiny)
již byla publikována, celý obor je však ve stavu rozvoje a na nové informace využitelné
i v akvaristice si budeme muset ještě počkat. Nicméně současný stav vědění dává na základní
otázky pěstování ponořených vodních rostlin dostatek odpovědí. Otázkou však je, zda jich
dokážeme v běžné praxi využít.
Řadu zajímavých souvislostí i množství odkazů na vědecké práce je možno nalézt v publikaci
Lubomíra Nátra.
Autoři níže uvedeného článku si kladli za cíl ověřit, který z faktorů - zda světlo či
koncentrace CO2 je tím hlavním limitujícím faktorem růstu ponořených vodních
rostlin.
V časopise AQUA PLANTA 4-2000 publikovali
v (německém překladu)
"Wechselwirkungen zwischen CO2 und Licht
stimulieren das Wachstum aquatischer Pflanzen"
Ole Pedersen, Troels Andersen
(University of Copenhagen) a Claus Christensen (Tropica Aquarium Plants)
výsledky provedeného měření závislosti intenzity růstu trhutky plovoucí
(Riccia fluitans) na dvou faktorech - intenzitě osvětlení a koncentraci CO2
ve vodě. Ačkoli výsledky získané tímto měřením pravděpodobně nelze mechanicky přenést na všechny
druhy ponořených vodních rostlin (*), jsou velmi názorným příkladem interakcí dvou základních
růstových faktorů v podmínkách blízkých poměrům v našich akváriích. Základní souvislosti
vyplývající z jejich měření jsou uvedeny dále.
(*) Trhutka (Riccia fluitans) je k takovým pokusům
mimořádně vhodná (pokrývá rovnoměrně rovinnou plochu - hladinu - v pokusné nádrži, její
vzdálenost od zdroje umělého osvětlení je snadno definovatelná a měřitelná stejně jako intenzita
dopadajícího světelného (FAR - fotosynteticky aktivní radiace) záření, vrstva vodního sloupce
snižující intenzitu osvětlení je zanedbatelná, snadno lze rostlinnou biomasu z nádrže vyjmout
bez poškození a zvážit pravidelné přírůstky zelené hmoty, příjem živin je plně zajištěn
z vodního sloupce - trhutka nemá vyvinut kořenový systém - vlastnosti této rostlinky umožňují
snadnou kontrolu výsledků opakovaným měřením - což o mnoha dalších druzích rostlin používaných
v akvaristice říci nemůžeme). Malým otazníkem je na druhé straně kontakt části porostu
s hladinou a vzduchem nad hladinou, odkud může přijímat CO2 i při jeho deficitu
ve vodě (a zkreslovat tím částečně výsledky měření). Je pravděpodobné, že podrobným rozborem
metodiky celého pokusu bychom odhalili i další výhody (zjednodušující měření), proč si autoři
vybrali právě tuto rostlinku.
Hlavním cílem již zmíněných autorů pokusu bylo ověřit, která z obou sledovaných
složek (světlo, CO2 ) má větší význam jako >>limitující faktor<< z pohledu
omezení intenzity fotosyntézy (růstu rostlin) při nasycení ostatními živinami. Chtěli také
ověřit, zda vyšší intenzita světla může (do jisté míry) kompenzovat nedostatek CO2
ve vodě a naopak, tedy zda dostatek CO2 ve vodě může zmírnit požadavky na intenzitu
osvětlení - t.j. zda se mohou světlo a CO2 vzájemně "zastupovat" při
nedostatku jednoho z nich, resp. který faktor má tuto schopnost vyšší.
Silné osvětlení odpovídá velmi intenzivně ozářeným akváriím - přírodní (málo tlumená)
intenzita světla,
slabé osvětlení pak akváriím se "standardním" ozářením
(pro účely pozorování akvária a života v něm - bez pěstitelských nároků). Střední hodnota
pak dobře osvětleným rostlinovým nádržím.
Mnoho CO2 ve vodě odpovídá přibližně 40 mg CO2 / litr
(cca 100-násobek nasycené úrovně ze vzduchu - viz
plyny ve vodě),
což je hodnota, která se v přírodě vyskytuje (dle údajů autorů) ve vodách zásobovaných
podzemními prameny bohatými na CO2 a pro akvária je doporučována jako hodnota
maximální (nebezpečí otravy ryb v akváriu).
Málo CO2 ve vodě pak
odpovídá pouhým desetinám mg CO2 / litr, což odpovídá přirozeným hodnotám.
(Jednotky [mg/l] a [lux] byly použity pro popularizaci, ve vědeckých pramenech
jsou uváděny údaje v [µmol/mol nebo mmol/mol] pro koncentrace v roztoku
a [W/m2] resp. [µmol [kvant]/m2. s]
pro intenzitu osvětlení resp. ozářenost.)
Základní schéma pokusu uvádí tabulka č. 1:
Tabulka 1
Závislost přírůstků rostlinné biomasy trhutky Riccia fluitans
na dvou proměnných faktorech

[denní přírůstky v %]
Z tabulky i grafu je zřejmé, že zvyšováním dodávky světla i CO2 směrem
k nasycené hodnotě dochází k výraznému (několikanásobnému) zvýšení přírůstků rostlinné biomasy
trhutky (Riccia fluitans). Zajímavé je, že několikanásobné zvýšení
koncentrace CO2 směrem k nasycené hodnotě u nízkých intenzit osvětlení nevyvolá
odpovídající zvýšení přírůstků zelené hmoty.
Současně vysoká dodávka světla i CO2 však vyvolává vysoké přírůstky biomasy.
Graf č. 3 znázorňuje, jak se 1 gram Riccia fluitans v uvedených světelných
a CO2 poměrech vyvíjel v průběhu dvou týdnů.
Graf 3
Graf závislosti přírůstků rostlinné biomasy trhutky (Riccia fluitans)
pro různé kombinace proměnných faktorů
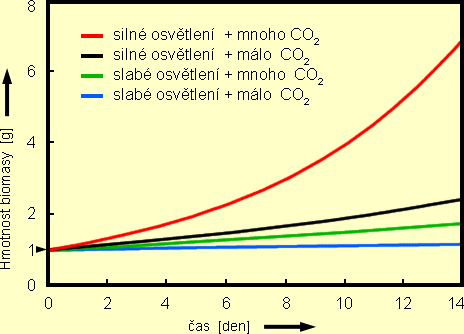
Z průběhu jednotlivých křivek vyplývá, že rozíly mezi různými kombinacemi jsou značné.
Pozoruhodná je intenzita růstu biomasy u současného intenzívního osvětlení,
přísunu CO2 a nasycení ostatními minerálními živinami. Naopak růst při malé intenzitě
světla a současném nedostatku CO2 jen málo přesahuje schopnost rostliny udržovat
"prostou reprodukci" vlastní zelené hmoty.
Přehled výsledků provedeného pokusu je zřejmý z následujícího obrázku. Při nízké
úrovni ozáření a množství dostupného CO2 nemá rostlina dostatek energie k optimalizaci
množství chlorofylu nebo enzymů. Zvýšením množství záření (FAR) nebo CO2 rostlina
uvolní část energie nutné pro využití původně nízké dodávky deficitního faktoru a tato může být
přesunuta k efektivnějšímu využití faktoru druhého, který zůstává nadále na nízké úrovni.
Zde autoři poukazují na podobnost s výsledky měření (pokusů) prováděných na jiných vodních
rostlinách (např. Elodea canadensis a Callitriche sp.) a vyvozují předpoklad,
že u většiny ponořených vodních rostlin budou naměřené závislosti podobné, jako v již
provedených výzkumech.
Obr. 1
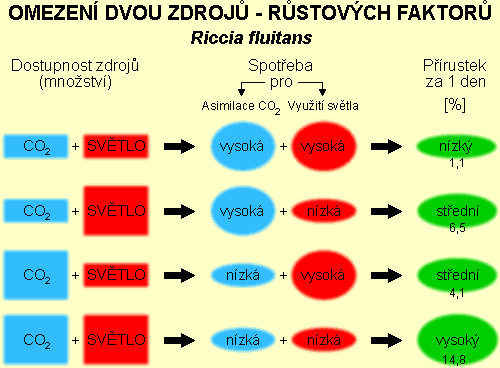
Z provedených pokusů a citovaných vědeckých prací podle autorů také vyplývá, že
Liebigův zákon minima (nedostatek jediného - limitujícího faktoru - způsobí omezení
růstu rostliny) neplatí za všech okolností zcela jednoznačně, že některé faktory (v tomto
případě FAR - světlo a CO2) se do určité míry mohou vzájemně kompenzovat - nedostatek
jednoho může být částečně kompenzován zvýšeným množstvím faktoru druhého.
Závěrem článku autoři poukazují na obtížnou ovladatelnost jednotlivých faktorů ve
složitém komplexu vztahů v akváriu. V nádržích s rybami doporučují využívat hnojiv bez obsahu
dusíku a fosforu. Konstatují obtížnost kontroly železa, manganu, draslíku a ostatních stopových
prvků - jejichž obsah závisí na materiálu použitém jako substrát dna.
Při řešení problémů růstu rostlin doporučují jako první krok (před přijetím dalších
opatření) nejdříve zvýšit koncentraci CO2 v akváriu. Pokud je v akváriu právě
CO2 limitujícím faktorem růstu, mělo by zvýšení jeho koncentrace přinést
viditelné zlepšení.
Autoři závěrem uvádějí seznam citovaných prací:
Andersen (1999): Interactions between light and inorganic carbon
stimulate the growth of Riccia fluitans L.
Report from The Freshwarer Biological Laboratory,
University of Copenhagen (v dánštině).
Maberley (1983): The interdependence of photon irradiance and free carbon dioxide
or bicarbonate concentrations on the photosynthetic compensation points of freshwater plants.
New Phytologist 93: 1-12.
Maberley (1985): Photosynthesis by Fontinalis antipyretica. I. Interaction between
photon irradiance, concentration of carbon dioxide and temperature.
New Phytologist 100: 127-140.
Madson (1993): Growth and photosynthetic acclimation by Ranunculus aquatilis L.
in response to inorganic carbon availability.
New Phytologist 125: 707-715.
Madsen and Sand-Jensen (1994): The interactive effect of light and inorganic carbon
on aquatic plants growth.
Plant, Cell and Enviroment 17: 955-962.
V textu byly m.j. použity údaje publikované v časopise AQUA PLANTA 4 - 2000:
Ole Pedersen*, Claus Christensen** a Troels Andersen*** (2000):
Wechselwirkungen zwischen CO2 und Licht stimulieren das Wachstum aquatischer
Pflanzen
(Proměnné vlivy CO2 a světla stimulují růst ponořených vodních rostlin)
*) University of Copenhagen,
opedersen@zi.ku.dk
**) Tropica Aquarium Plants,
clc@tropica.dk
***) University of Copenhagen,
tandersen@zi.ku.dk
Originál původního textu v angličtině je dostupný zde:
[1],
[2],
[3],
[4]...
Vladimír Pelikán